Introduction
어류 및 갑각류의 종묘생산 연구가 활발해짐에 따라 유생기의 먹이로서 Artemia spp. (Brine shrimp)의 내구란에 대한 수요가 점차 많아지고 있다.
·Brine shrimp는 어류와 갑각류 유생기의 초기 먹이생물로서 적당 (DEMPSER, 1953) ·방사선과 독물등의 생물 검정재료로서 사용 (MICHALE, 1956)
·염전에서 번식하는 식물 Plankton을 섭취하여 祖 의 착색을 방지 (RACKETT, 1812)
·식용으로 사용 (ABONYI, 1915)
Brine shrimp의 nauplius가 종묘생산용 먹이로서 좋은 이유는 부화직후 영양가가 높을뿐만 아니라 내구란의 보존이 용이하고 수시로 부화시킬수 있다는 것인데, 아직까지 채란보존과 부화등에 대한 지식은 충분치 못하다고 한다. (二材, 1968)
여기에서는 Brine shrimp의 생태 및 생리에 대한 기초적인면, 즉, 분류, 분포, 생식, 부화, 성장, 섭이, 호흡 및 삼투압 조절등에 관해서 알아보기로 한다.
1.分類
Brine shrimp는 함수산 하등갑각류로서 분류학상 Arthropoda, Crustacea, Branchiopoda, Anostraca, Artemia科에 속한다(LINDER, 1941)
Anostraca은 male의 제 2촉각(DADAY, 1910) 또는 생식기(陰經)의 형태(LINDER, 1941)에 따라서 科가 분류되는데 female에게서는 볼수 없으므로 種에 대한 조사가 곤란하다.
또한, 함수화 기수에 서식하는 Anostraca의 종류에는 本種이외에 Brinchinecta ferox, Brinchinectella salina, Chilocephalus stagnalis, Paratemia contracta, P. serventyi, P.zietziana등, 6종이 있다고 한다.
LINDER(1941)에 의하면 Artemia는 다행히 female의 육아낭(育房)에 특징이 있고 배쪽에 하나의 을 가지고 있는데 이와 같은 형태는 Anostraca중에서 Tanymustix affinis, T. stagnalis 에 한정되어 있다고 한다.
T. affinis, T. stagnalis는 크기에 있어서 Artemia와 비슷했지만 꼬리부기형태는 다르다. (그림1) 즉, Tanymustix spp.은 꼬리분기가 칼과 같은 형태로서 가늘고 길지만 Artemia 에서는 그렇지 않다는 것을 알 수 있다.
또, Artemia 변종에 대한 분류는 꼬리분기의 형태로서 (ABONYI, 1915) 나누어지며, 분기형태는 염분도에 의해서 변화할수 있다고 한다.
2. 分布
Brine shrimp는 양극지방이외의 Greenland를 포함한 전세계에 분포한다고 알려져 있으며, 50 strains 있다고 한다. (Handbook of Mariculture, 1983)
Brine shrimp는 완전한 담수에서 하루동안 살수 있으며, 염분도 35‰, 수온 25℃가 표준상태이지만, 염분도 5∼150‰, 수온 6∼35℃의 범위와 용존산소 1ppm이하에서 생존할수 있는 광염성, 광온성이다. (Handbook of Mariculture, 1983)
3. 生殖
Brine shrimp는 자웅이체로서 암수가 항상 있는 種은 유성생식을 하며, female만 있는 경우에는 처녀생식을 하다가, male이 출현하면 쌍방의 생식에 의해서 번식한다.(ARIOM, 1911)
Brine shrimp 출산에는 어미의 育房에 있는 생식공에서 nauplius로 출산하는 난태생과, 내구란을 방란하는 난생의 2가지 양식이 있다.
male의 제 2촉각은 발달하여 把握器(clasper)로 되어있고 제 12, 13체절은 생식체절로 불리워지고 있으며, male은 방추상의 발달되어 있지 않다.
female의 난소는 장이 좌우로 뒤쪽에 줄지어 있고, 앞부분에 있는 수란관을 통해서 育房에서 산란한다. 교미는 성숙한 male의 제 2촉각으로 female의 育房의 앞부분을 잡고 배쪽을 위쪽으로 하며 같이 유영하고, 때에 따라서는 복부를 많이 휘어서 교미하는데, 교미는 1분이내에 끝난다.
출산은 유성생식에 의해 매회 교미가 필요한데 (BOWEN, 1962) 수온 28℃에서 교미후 50∼80시간 사이에 출산하며 1회 산란수(卵生인 경우)와 産仔수(난태생인 경우)는 2∼30개인데 많을 때에는 80∼150개 된다. (BOWEN, 1962)
출산간격은 28℃에서 2.5日이상이고 보통 5∼10회 출산을 한다.
4. 孵化
부화는 우선 바깥쪽의 두꺼운 난각과 얇은 막을 벗어 버리고 얇은 막과 같이 있는 또 하나의 막에 쌓여져서 nauplius의 頭部를 밑으로 하여 매달린다. 다음에 쌓여져 있는 막의 머리부분부터 째져서 nauplius가 헤엄쳐 나오는데 JENNINGS 와 WHI (1941)는 앞단계를 emerge(-nce), 나중의 단계를 hatch(-ing)이라 구별하고 있다.
전자는 삼투압에 의한 각의 파괴이고, 후자는 부화효소가 관여하는 것이라 생각된다.(石田壽老, 1948)
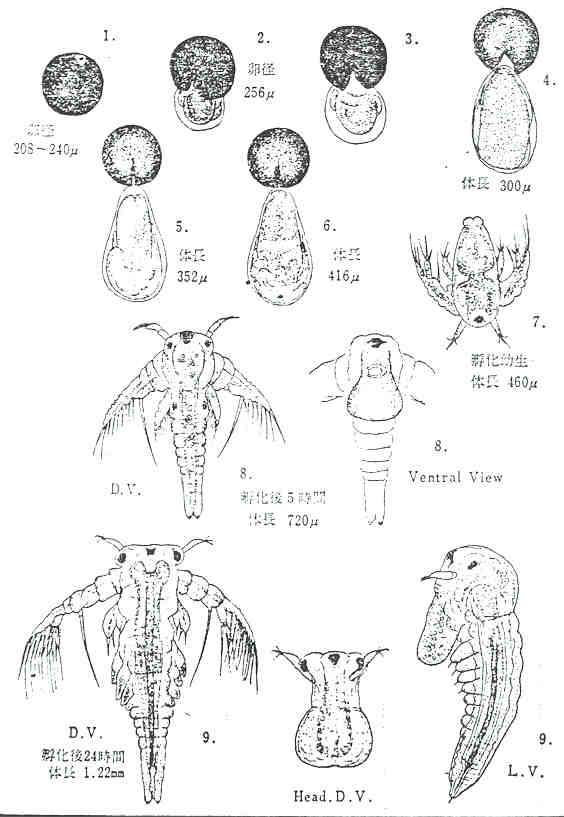
부화직후 유생은 체장 0.45㎜, 체폭 0.17㎜, 무게 0.01㎎정도가 된다.
부화조건에 대한 조사를 보면
·BOONE 와 BASS-BECKING (1931)은 수온에 대한 조사에서, California 産개체를 해수 (필자추정)에서 부화시켜 그림 4와 같은 결과를 얻었으며
·岡山縣수산시험장(1965)에서 戶內연안의 宇野産개체를 대항으로 한 실험에서도 같은 결과를 얻었다.
·BOONE와 BASS-BECKING (1931)은 염류조성의 영향에 대해서 조사한 결과 K+〉Ca+〉Mg++의 순으로 염화물의 옥성이 인정되는데 Na+이온은 그러한 것에 길항작용을 가진다고 하였다
이와 같은 이온은 emerge에는 영향이 없으나 hatch에서는 영향을 미치고 있으며, emerge는 증류수중에서도 볼 수 있다.
·JENNINGS와 WHITAKER (1941)는 California 産개체를 사용해서 해수를 기준으로 하여0∼56범위의 농도에서 부화시킨 결과 모두가 약 60% emerge되고, 부화율(hatch/emerge) 및 emerge와 hatch의 속도와 염분도의 관계는 그림 5와 같이 나타나고 7∼30범위의 해수에서는 hatch한 속도가 빠르다는 것을 알 수 있다.
5. 成長
Brine shrimp는 nauplius로 부화되어 뚜렷한 변태를 하지 않고 성체가 된다.
난생, 난태생을 불문하고 부화직후의 nauplius는 체장이 약 0.45㎜이고, 제 1 및 제 2 촉각을 가지며, 주로 제 2 촉각을 사용하여 유영한다.
HEATH(1924)에 의하면 Brine shrimp의 nauplius는 제 1令의 말기에 소화관이 완성되어 섭식을 시작한다고 하였으며, 그 후 탈피와 동시에 체절과 유영지의 수가 많아지고, 발달하여 체장 2∼3㎜에서 유영이 활발해지며 암수를 외부형태에 의해 구별할수 있게된다. (PROVASOLI and SHIRAISHI, 1959)
성장은 수온, 염분도, 먹이 등에 의하여 좌우되는데 28℃전후에서는 부화후 하루이내에 먹이를 먹기 시작하고, 8일째에는 riding하고 (교미도 같은날 한다고 생각됨) 11일째에 출산한 예가 있으나(寺本賢一郎, 1961) 일반적으로 부화후 2∼3주 사이에 출산한다. 성체는 체장 10∼15㎜이며, 그림같이 female이 최종적으로 약간 크게 된다.(GILCHRIST, 1960) 또 성체의 크기는 염분도가 높을수록 작아지는 경향이 있는데 성장속도는 염분도가 그다지 영향이 없다.(二村, 1967) 염분도에 따라서 형체가 변화한다고 하는데 BAID(1963)는 염분도가 높을수록 복부가 길게된다고 하였으나 그렇지 않다는 이론도 있다.(GILCHRIST, 1960) 체장과 체중의 관계는 그림 7과 같다.
·Brine shrimp의 일반 성분을 종합해 보면 표 1과 같이 되며 섭식개시후는 소화한 내용물이 전체에 대해서 약간 더 많은 함량을 차지하는데 그 영향을 고려해야 할 것이다.
6.攝餌
Brine shrimp는 protozoa, microalgae, Yeast, Bacteria 등을 먹는 잡식성이며 전형적인 tilter teeder 인데 때에 따라서는 scraping 한다.
규조류의 Chaetoceros 와 Skeletomema 등은 가장 좋은 먹이이며 Chlamydomomas sp.,해산 Chlorella 등도 Brine shrimp 의 먹이로서 적당하다.
섭이는 nauplius 시기에서는, 주로 제 2 촉각에서 모은 입자를 다시 大顎의 입술부분에 모아서 소화관에 밀어 넣는 방법으로 행해지며 (GAULD, 1959) 성체에서는 유영지의 복부 한가운데 線(腹部正中線)의 food grove로 모은 것을 다시 유영지의 내지기부(遊泳肢の 內肢基部)에 있는 강모(剛毛)에서 앞쪽으로 보내거나, 입술, 입부분의 부속지에 의해서 소화관에 밀어 넣는다.(CANNON, 1933)
유영지에 의해서 단위시간에 여과된 수량은 여수율 (여수량, fiterating rate, ㎖/尾/日)이고 실제로 섭취된 이료의 양(세포수)은 섭이율 (섭이속도, grazing rate, 세포수/尾/日)로 표시되며 양자는 명확히 구별되어야 한다.
여수율은 섭이 능력을 표시한 것인데 수온, 체장등에 의해서 현저한 차이가 있다.
·REEVE(1963)는 여러 가지 크기의 Brine shrimp의 여수율과 섭이율을 이료농도로 바꾸어서 측정하였는데 이에 의하면 저이료 농도에서 여수율은 일정하였고, 임계 농도 이상에서는 섭이율이 일정하여 여수율은 떨어진다고 하였다.
여수율과 체장의 관계는 그림 8과 같다. 최대 섭이율은 Chlamydomomas sp.(10∼15×6∼9u)을 이료로 할 때에 수온 28℃, 체장 5㎜의 Brine shrimp에서는 약 700만 세포/尾/日(二村, 1964)이고, 이러한 경우 유지식량(維持食量) 10∼30만 세포/尾/日(二村, 1963)에 비하며 현저하게 크다.
이와 같이 filter feeder는 일반적인 필요량에 비하여 아주 큰 섭이 능력을 가지고 있는 것 같다.
7.呼吸
체장과 호흡량과의 관계는 von BERTALANFFY 와 KRYWIENCZK(1953)이 KIRK 의 장치로 측정하여 체장과의 관계를 구하였고 CONOUER(1960)은 WINKLER 법으로서 Brine shrimp 의 전 질소량, 건중량에 대한 관계를 조사하였다. 호흡량과 체장, 수온의 관계는 그림 9과 같다.
·GILCHRIST(1956)에 의하면 性에 의한 차이는 일반적으로 인정되어 있지 않은데 염분도가 높아짐으로써 fomale이 male 보다 산소를 많이 소비한다.
그리고 naupius의 호흡량은 밝은 곳에서나 어두운 곳에서나 변화가 없다고 한다.(ELIASSEN. 1952)
용존 산소 농도의 영향에 대해서 GILCHRIST (1956)가 헤모그로빈의 기능과 관련하여 조사한 바에 의하면 21∼22℃의 해수중에서 헤모그로빈을 가지고 있지 않은 개체는 산소포화도 20% 정도에서 2日후 반수 이상 사망했는데 헤모그로빈을 가진 개체에서는 사망률이 없었다고 한다.
또 염분도 190‰ 에서는 포화도 80%(1.6㎖O2/L)이상이 되면 호흡량은 대개 일정한데 헤모그로빈을 가지고 있는 것은 헤모그로빈을 CO에서 저해하는 것 보다 많이 호흡하며, 양자의 차이는 포화도 53%(1.05㎖O2/L) 이하에서는 많이 없어진다고 한다. 그렇지만 Brine shrimp의 서식염분 범위가 넓고, 같은 산소분압에서도(포화도) 염분도가 현저하게 다른 농도(㎖O2/L)에서는 많은 차이가 있는데 앞으로 더욱 연구 되어져야 한다.
·내구란이 부화할 때 사용되는 물질대사에 대한 산소소비량의 측정예를 보면 그림 10과 같다.
8.渗透壓 調節
광범한 염분범위의 함수에 서식하며 어느정도의 급격한 염분변화에 견딜수 있는 Brine shrimp는 큰 삼투압 조절 능력을 가지고 있다. CROGHAN(1958)이 환경수의 감투압과 체액의 삼투압을 빙점 강하법에서 측정하였는데 이것에 상당한 Nacl 용액 농도로 환산하면 그림 11과 같이 된다.
9. 要約
1. Brine shrimp는 Arthropoda, Crustacea, Branchiopoda, Anostraca, Artemia 科에 속한다.
2. Brine shrimp는 완전한 담수에서 하루동안 살수 있으며 염분도 35‰, 수온 25℃가 표준상태이지만 marvelously euryhaline, eurythermal, 그리고 대표적인 euroxybiont 이다.
3. Brine shrimp는 자웅이체로서 유성생식을 하지만 female만 있는 경우에는 처녀생식을 한다.
4. 부화의 과정은 emerge(-nce)와 hatch(-ing)의 두 단계로 구별된다.
5. Brine shrimp는 부화후 2∼3주 사이 출산하며 성체의 크기는 10∼15㎜이며 수명은 2∼3개월이다.
6. Brine shrimp는 전형적인 filter feeder 이며 때에 따라서는 scraping 하기도 하며 protozoa, microalage, Bacteria, Yeast 등을 먹는 잡식성이다.
7. nauplius의 호흡량은 明暗과는 관계가 없다.
*배양장에서는 보통 고밀도로 대량 부화하게 되므로 최상의 부화를 위해서는 아래의 주요 지표를 잘 관리해야 한다.
*특히 부화시간이 길어지면 일찍 부화된 유생은 자체의 함유 영양가가 점점 떨어지게 되므로, 전 개체의 관점에서 부화방법과 먹이
급여시간이 일정하게 표준화되어야 한다.
즉, 먹이를 효율적으로 주기 위해서는 한정된 짧은 시간애에 최대한 많은 갓부화한
노우플리우스를 생산할 수 있는 최적 부화조건을 항시 유지해야 한다.
(1) 온도
28℃를 유지한다. 보통 아르테미아의 부화온도는 25∼30℃이며, 25℃ 이하에서는 부화가 지연되고 33℃
이상에서는 대사가 멈추게 되므로, 에너지 손실없는 최상의 갓부화한 노우플리우스를 많이 얻기 위해서는 적정온도인 25∼28℃를 일정하게
유지해야 한다.
(2)염도 및 pH
사용에 편리하기 때문에 보통 자연해수를 그대로 사용한다. 염도는 28∼34‰ 정도가 적당하다. 하지만 pH는 부화기간중에 8 이상을
유지하는 것이 매우 중요하다. 부화중에 pH가 떨어지게 되므로 최상의 부화를 위해서는 시작 전에 NaHCO3을 첨가하여 pH가 8.5이상
되도록 한다.
부득이 pH가 낮은 물로 부화를 해야 할 경우 아르테미아 알의 밀도를 낮추는 것이 좋다.
염도 28∼34‰은
물 1.000cc에 염분이 28∼34g 들어 있는 해수농도이다.
(3)에어레이션(aeration)
최상의 부화를 위해서는 20mg/ℓ 이상의 산소 용존량의 유지가 필요하다. 에어레이션은
산소공급 뿐만 아니라 알을 부유하게 하는데 중요하다. 가라앉은 알은 부화하지 않으므로 에어레이션이 골고루 충분하게 되도록
한다.
(4)밀도
대량 부화시 용기 상태. 에어레이션. 부화된 노우플리우스의 사멸 등을 감안하여 결정하여야 하나, 해수 1ℓ당
2g을 초과하지 않아야 한다. 우리 나라 여건으로는 1ℓ당 2g 정도가 적당하다고 여겨진다. 고밀도 부화시 거품의 발생에 유의해야
한다.
(5)광도
부화 시작 후 1시간 동안은 빛이 필수적이며, 부화기간 중에 수면 위로 1.000Lux의 빛을 일정하게 유지하는
것이 이상적이다.
(6)소독
알의 껍질에는 박테리아 등의 병원균이 많이 묻어 있어서 고수온하에 고밀도의 알이 들어 있는 부화용기에서 잘
번식하여 노우플리우승 옮기게 되므로 부화시작 전에 알과 용기의 소독이 필요하다. 소독이 어려운 여건에서는 수돗물에 알을 1시간 정도
담궜다가 125㎛ 정도의 망에 넣어 세척하면 병원균 전염을 최소화할 수 있다. 소독은 클로린(Chlo-rine) 계통의 소독제를
사용하는 것이 일반적이다.
(7)용기
대량 부화를 위해서는 바닥으로부터 에어레이션이 고루 퍼질 수 있는 투명한 원통형 용기가 좋으며, 수온과 광도를
일정하게 유지할 수 있는 장치가 구비되어야 한다.
(8)채집요령
*부화가 끝나 에어레이션을 끄면 부화되지 않은 알껍질은 수면 위로 뜨고, 부화된 노우플리우스는 바닥에 가라앉게
되므로 5∼10분 후부터 노우플리우스를 분리채집한다.
*주관성을 이용하여 용기의 윗부분을 어둡게 가리고 밑의 한 부분만을 밝게 빛을 쪼이면
그 곳으로 노우플리우스가 모이게 되어 쉽게 채집할 수 있다.
*채집시에 노우플리우스를 공기에 노출시키거나 조그만 용기에 밀집시키면
산소부족으로 활력을 잃게 되므로 항시 맑은 물이나 에어레이션을 공급하면서 채집하도록 해야 한다.
(9)먹이 주는 요령
①부화 용수에서 묻기 쉬운 글리세롤 찌꺼기 및 병원균 등을 없애기 위해 부화된 노우플리우스는
부화용기로부터 호스를 통해 물 속에 잠긴 플랑크톤망(100∼125㎛)에 모으고 깨끗한 물을 주수하면서 잘 세척한 후 치어탱크에 넣어주어야
한다. 새척시에 압력이 센 물이 직접 닿으면 노우플리우스가 상처를 입기 쉬우므로 조심해야 한다.
②부화된 노우플리우스는 전적으로
저장된 에너지로 살아가기 때문에 가장 에너지가 많은 상태(갓부화)로 먹이도록 하여야 한다. 보통 첫 유생단계에서 두 번째 단계로 변태하면
초기 에너지량의 27% 정도가 감소할 뿐만 아니라 먹이효율도 떨어진다.
작업형편상 일시에 대량 부화했지만 여러 차례에 나누어 주어야
할 경우 ㎖당 15,000개체 이하의 밀도로 에어레이션하면서 0∼4℃ 냉장고에 보관하거나 비닐봉지에 싼 얼음을 넣어서 0∼10℃
정도의 수온을 만들어 보관하였다가 먹이면 영양손실과 크기가 커지는 것을 최소화할 수 있다.
③ 아르테미아 유생이 활발하게 유영하여야
치어가 잘 먹으므로 유생의 활력이 좋도록 유지해야 한다.
④ 대부분 어종의 자어는 주로 시각에 의해 먹이생물을 체포하므로 자어탱크의
벽면색깔(유생과 대비되는 짙은 색이나 검정색)과 광도(밝기)를 고려하는 것이 먹이를 찾아 먹는데 도움이 된다.
5. 물벼룩의 발생
· 물벼룩 무리는 봄이 되면 전년도 가을에 못바닥에 떨어져 있다가 겨울을 지난 알에서 부화한다. 이들은 모두 암컷들 뿐이고, 수컷없이 단위생식을 하여 자충을 산출한다. 이들 자충은 수온·수질·먹이 등의 조건이 좋으면 6∼7일만에 어미체가 되어 다시 단위생식(單爲生殖=單性生殖)을 하며, 3∼4일 마다 계속 산란한다.
· 이와 같이 단위생식을 거듭하게 되면 급격히 증식이 되지만, 수온이 내려가거나 먹이가 부족하게 되면 수컷이 나타나서 유성생식(有性生殖)을 하여 다시 내구란(耐久卵 또는 )을 산출하게 되어 번식이 증지된다.
· 물벼룩의 크기는 종류에 따라 다르지만, 성장하면 1∼2㎜가 된다. 이러한 것을 부화된 자어가 먹기에는 약간 큰 상태이다. 즉, 단위생식에 의해서 산출된 자층은 0.1∼0.5㎜이며, 이것은 초기 자어의 먹이로서 알맞다.
· 물벼룩을 발생시키려면 부화하기 15일 전에 사육지의 물을 빼고, 150∼300g/㎡의 생석회를 살포한 후 1일 이상 경과한 후에 한 번쯤 새물로 잔류석회를 용해시켜 배수한 후 유기질비료 또는 무기질비료를 시비한다. 유기질 비료는 분해가 늦으나 효과가 오래 지속된다.
· 시비방법은 아래 표에서와 같이 보통 닭똥·퇴비 등을 혼합하여 1㎏/㎡ 비율로 뿌려준다. 그리고 무기질 비료는 복합비료를
3.3㎡당 40∼50g정도(300평당 유안 10kg, 중과석 5kg, 염화칼리 1kg의 비율)로 뿌려주며, 이것을 5일 간격으로 3회
계속 뿌려준다. 첫 번 시
비 후 약 30∼50㎝ 정도의 깊이가 되도록 물을 넣어주면, 7∼10일 후에는 물이 적갈색에서 녹갈색으로
변하게 되면서 이때부터 물벼룩이 많이 발생된다.
6. 실지렁이의 소독 처리방법
· 실지렁이를 구입한후 유수처리로 측양하여 장내 세균 및 뻘 등 불순물이 완전히 배출되도록
한다. 먹일 때는 Furasol로 소독 처리하고, 영양제를 투여한 후에 치어에게 준다.